Результат интеллектуальной деятельности: РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pcDNA4-Apo-2NLS2, НЕСУЩАЯ СИНТЕТИЧЕСКИЙ ГЕН БЕЛКА АПОПТИНА, ИНДУЦИРУЮЩЕГО p53 НЕЗАВИСИМЫЙ АПОПТОЗ ОПУХОЛЕВЫХ КЛЕТОК
Вид РИД
Изобретение
Изобретение относится к рекомбинантной плазмидной ДНК со встроенным геном белка апоптина, индуцирующего апоптоз неопластических клеток человека, перспективного для использования в качестве противоопухолевого терапевтического трансгена, и может быть использовано в молекулярной биологии и микробиологии.
Несмотря на успехи в терапии злокачественных новообразований, прогресс, достигнутый с применением классических методов лечения, таких как хирургия, радио- и химиотерапия, в настоящее время представляется достигшим своего предела. Летальность до года с момента установления диагноза и начала лечения составляет 35-37%, а 5-летняя выживаемость пациентов не достигает 50%. Существенным недостатком химио- и радиотерапии является их низкий терапевтический индекс, из-за чего увеличение дозы или комбинирование подходов для преодоления устойчивости или усиления разрушения раковых клеток сопровождается токсичностью и повреждением нормальных тканей. Поэтому потребность в разработке новых эффективных и высокоспецифичных средств терапии онкологических заболеваний чрезвычайно актуальна.
Апоптин является одним из неструктурных белков вируса анемии цыплят (VP3), который отвечает за гибель клеток при вирусной инфекции [1-3]. Апоптин способен индуцировать апоптоз широкого спектра раковых и трансформированных клеток, но не нормальных клеток 4, 5]. Это белок длиной 121 аминокислот (14 кД). Апоптоз, вызываемый апоптином, не зависит от р53 и не подавляется экспрессией антиапоптозного белка Bcl-2. Эти свойства делают апоптин привлекательным кандидатом в терапевтические белки, которые могли бы усилить онколитические свойства вирусов.
Механизм действия апоптина до конца не выяснен. Однако известно, что в опухолевых клетках апоптин локализуется преимущественно в ядре, в то время как в нормальных клетках он расположен в цитоплазме. Впрочем, возможно, что ядерная локализация - причина, а не результат индукции апоптоза.
Аминокислотная последовательность апоптина содержит два основных сигнальных участка: NLS (NLS1: аминокислоты 82-88, и NLS2: аминокислоты 111-121) - отвечающий за ядерную локализацию апоптина (его отсутствие приводит к диффузному распределению апоптина по клетке) и NES - последовательность необходимая для выхода апоптина из ядра (в опухолевых клетках), На фиг.1 представлена первичная структура апоптина. Ключевые домены апоптина выделены цветом. NLS - сигнал ядерной локализации; NES - сигнал предполагаемого ядерного экспорта.
Известен следующий механизм действия апоптина в опухолевых клетках [6]. При попадании в клетку апоптин связывается с регуляторной субъединицей PI3-киназы, вызывая ее конститутивную активацию, которая ведет к продолжительной активации и ядерной транслокации киназы Akt. Находясь в ядре, Akt активирует киназу клеточного цикла CDK2 посредством как прямого фосфорилирования, так и непрямым способом, включающим индукцию протеасомного разрушения ингибитора CDK белка p27/Kip1. Активированная киназа CDK2 фосфорилирует апоптин по Thr-108 и тем самым способствует его ядерной локализации в раковых клетках. В ядре апоптин может связываться с рядом белков-партнеров, таких как DEDAF, Nmi, APC/C и PML. Пока невыясненным образом апоптин способствует фосфорилированию белка Nur77 и его экспорту из ядра. Известно, что в цитоплазме Nur77 модулирует функцию Bcl-2-подобных белков, превращая их из анти-апоптотических в про-апоптатические молекулы. Эти и, возможно, другие события способны инициировать апоптоз через митохондриальный путь активации каспаз.
Известно несколько рекомбинантных плазмидных ДНК для транзиентной экспрессии со встроенным геном белка апоптина, а также вирусных векторов (рекомбинанты аденовируса, вируса болезни Ньюкасла, лентивирусов), его экспрессирующих [6, 7, 8], обладающих избирательной онколитической активностью обусловленной селективным апоптоз индуцирующим воздействием на опухолевые клетки.
Наиболее близкой к заявляемой плазмидной ДНК является эукариотическая экспрессирующая векторная плазмидная ДНК-pcDNA3/Apoptin (прототип), способная индуцировать апоптоз неопластических клеток человека [9]. Описана плазмидная ДНК Ad-CMV-Apoptin, с использованием которой и аденовирусной векторной системы Adeno-X Expression System (BD Biosciences Clontech) были получены рекомбинантные аденовирусы, экспрессирующие ген белка апоптина, обладающие онколитической активностью in vitro и in vivo [10].
Однако известные аналоги и прототип имеют недостаточную избирательную апоптоз индуцирующую активность эксрессируемого модифицированного гена белка апоптина в отношении опухолевых клеток.
Техническим результатом является повышение избирательной апоптоз индуцирующей активности эксрессируемого модифицированного гена белка апоптина в отношении опухолевых клеток.
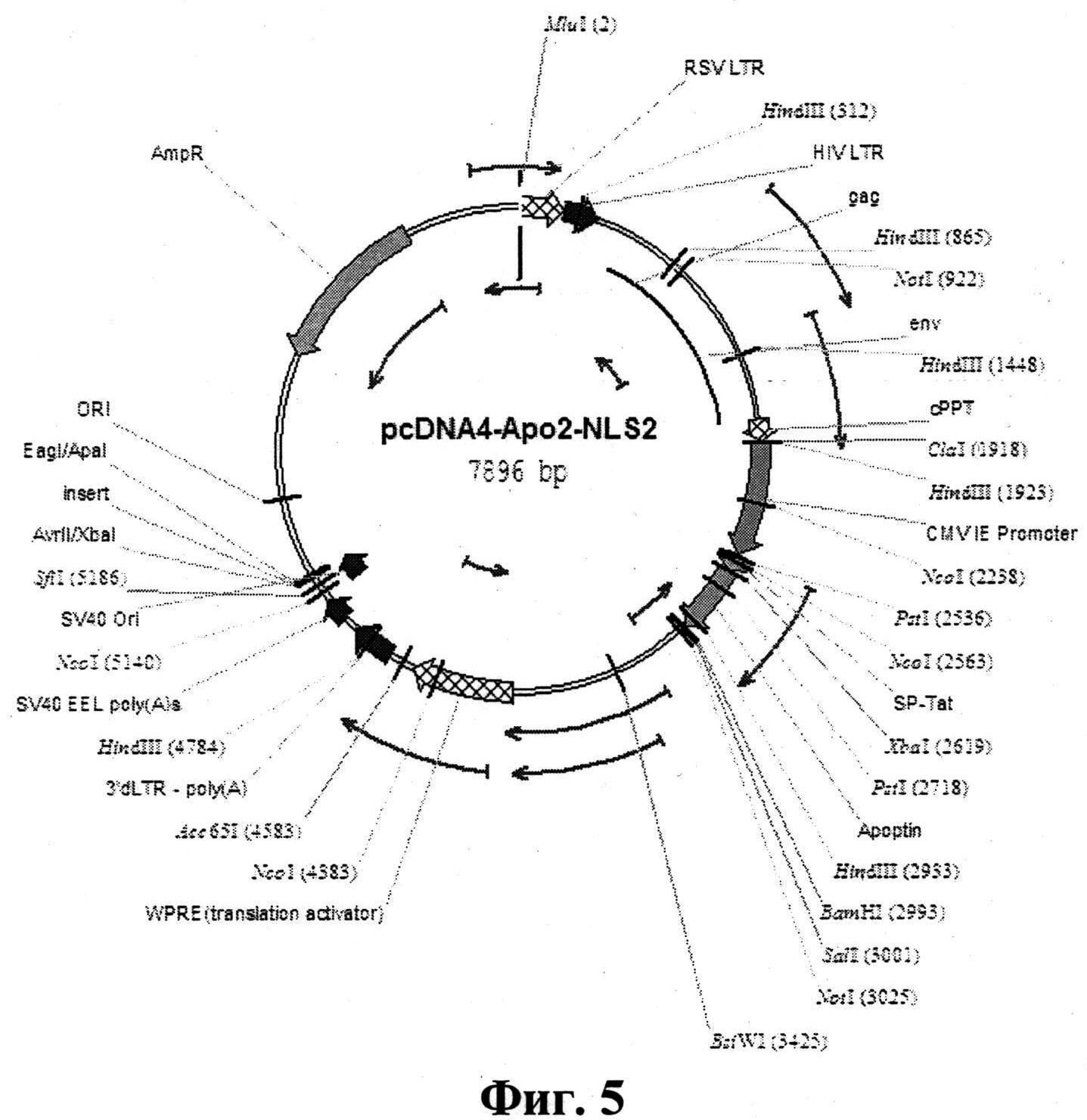
Указанный технический результат достигается тем, что синтезирована рекомбинантная плазмидная ДНК pcDNA4-Apo-2NLS2, несущая синтетический ген белка апоптина с удвоенным сайтом NLS2, избирательно индуцирующего р53 независимый апоптоз неопластических опухолевых клеток человека, с нуклеотидной последовательностью SEQ ID NO1 размером 393 п.н.,, представленной на фиг. 4, имеющая молекулярную массу 5,13 мДа, размер 7896 п.н. и содержащая в соответствии с физической и генетической картой, приведенной на фиг. 5:
- оперон размером 1071 п.н., состоящий из ранне-позднего промотора CMV, лидерного фрагмента, последовательности пенетрирующего пептида белка ТАТ ВИЧ, гена апоптина вируса анемии цыплят и последовательности эпитопа Flag;
- промотор CMV 232-819 п.н.;
- Т7 промотор 863-882 п.н.;
- Xpres праймер 1112-1130 п.н.;
- ТОРО сайт клонирования 1184 п.н.;
- генетические маркеры:
ген устойчивости Zeocin;
ген b-лактамазы, определяющий устойчивость к ампициллину;
- уникальные сайты рестрикции:
ClaI (1918); XbaI (2619); BamHI (2993):
SalI (3001); BstWI (3425).
Основным отличием предлагаемой рекомбинантной плазмидной ДНК от вышеприведенных конструкций (аналогов) является то, что встроенная в плазмиду pcDNA4 генная кассета, несущая последовательность гена белка апоптина, содержит удвоенный сигнальный участок NLS2, что предполагает возможность усиления избирательной апоптоз индуцирующей активности эксрессируемого модифицированного гена белка апоптина в отношении опухолевых клеток по сравнению природным геном апоптина за счет увеличения его способности к внутриядерной локализации в неопластических клетках.
Заявляемая плазмидная ДНК перспективна для разработки на ее основе онколитического препарата для генной терапии злокачественных новообразований человека, а так же для конструирования с ее использованием вирусных векторных систем с целью получения высокоэффективных и безопасных противоопухолевых онколитических вирусных препаратов.
Конструирование плазмидной ДНК pcDNA4-Apo-2NLS2.
Синтез гена белка апоптина с удвоенным сайтом NLS2 осуществляли методом элонгации праймеров в ПЦР. С этой целью по прототипной последовательности гена белка VP3 (апоптин) вируса анемии цыплят (GenBank Р54094) были рассчитаны, химически синтезированы и использованы следующие праймеры, представленные на фиг.2.
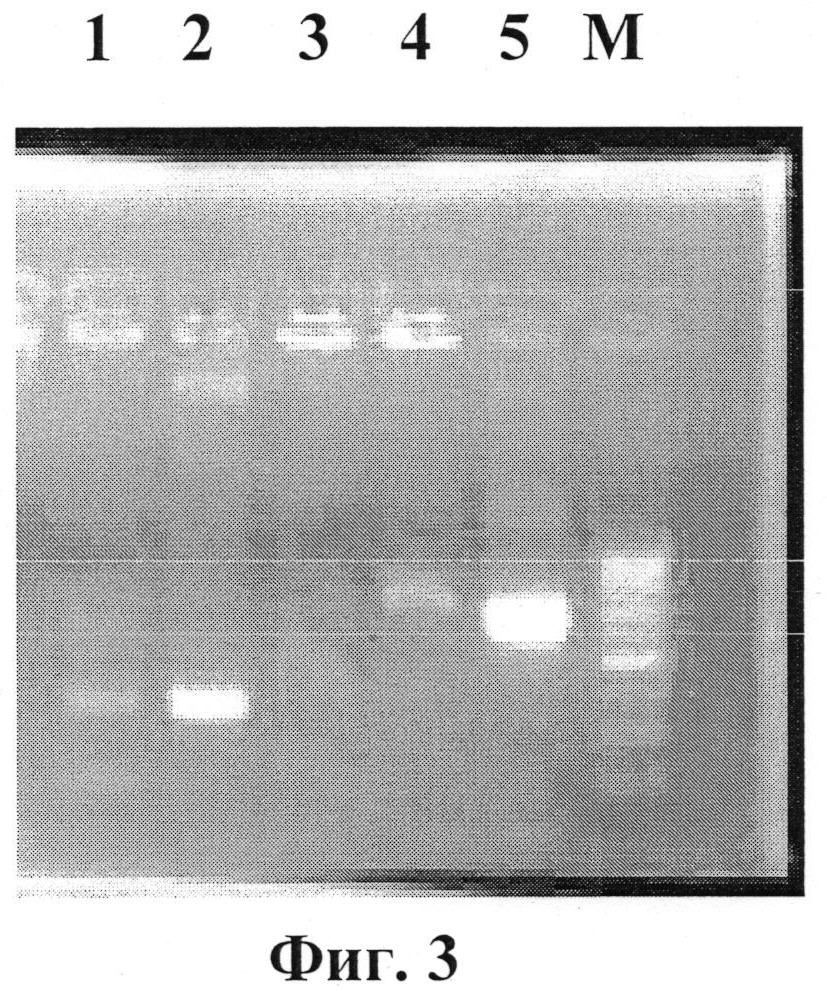
На фиг.3 приведена электрофореграмма синтезированного фрагмента ДНК, содержащего ген белка апоптина. Длина фрагмента соответствует ожидаемой (471 п.н.) и включает ген белка апоптина (393 п.н.) и последовательности ДНК (78 п.н.), комплиментарные аналитическим праймерам и фланкирующие целевую последовательность. Электрофорез в 2% агарозном геле фрагмента ДНК 471 п.н. содержащего ген белка апоптина (дорожки 1 и 2). М - маркер молекулярных весов.
Нуклеотидная последовательность SEQ ID NO1 синтезированного гена белка апоптина (393 п.н.) определена секвенированием и приведена на фиг.4.
Синтезированная нуклеотидная последовательность была клонирована в челночную плазмиду для транзиентной экспрессии pcDNA4TOPO (Invitrogen) по сайтам рестрикции XbaI-BamHI, в результате чего была получена рекомбинантная плазмидная ДНК pcDNA4-Apo-2NLS2, несущая синтетический ген белка апоптина с удвоенным сайтом NLS2, избирательно индуцирующего р53 независимый апоптоз неопластических опухолевых клеток человека, с нуклеотидной последовательностью SEQ ID NO1 размером 393 п.н., представленной на фиг.4, имеющая молекулярную массу 5,13 мДа, размер 7896 п.н. и содержащая в соответствии с физической и генетической картой, приведенной на фиг.5, оперон размером 1071 п.н., состоящий из ранне-позднего промотора CMV, лидерного фрагмента, последовательности пенетрирующего пептида белка ТАТ ВИЧ, гена апоптина вируса анемии цыплят и последовательности эпитопа Flag; промотор CMV 232-819 п.н.; Т7 промотор 863-882 п.н.; Xpres праймер 1112-1130 п.н.; - ТОРО сайт клонирования 1184 п.н.; генетические маркеры: ген устойчивости Zeocin; ген b-лактамазы, определяющий устойчивость к ампициллину; уникальные сайты рестрикции: ClaI (1918); XbaI (2619); BamHI (2993): SalI (3001); BstWI (3425).
Для сравнения была получена плазмидная ДНК pcDNA4-Apo (аналог), несущая генетически не модифицированный ген белка апоптина, которую в дальнейшем использовали в качестве контрольной. Правильность структуры сконструированных плазмид была подтверждена рестриктным картированием и секвенированием.
На фиг.5 приведена физическая и генетическая карта заявляемой плазмиды pcDNA4-Apo-2NLS2, несущей генетически модифицированный ген белка апоптина, а на фиг. 6 - структура этой рекомбинантная плазмиды. На Фиг. 7 представлена аминокислотная последовательность белка апоптина, экспрессируемого рекомбинантными плазмидами pcDNA4-Apo-2NLS2 и pcDNA4-Apo.
Исследование апоптоз индуцирующей активности рекомбинантной плазмидной ДНК pcDNA4-Apo-2NLS2 при трансфекции опухолевых и нормальных клеток человека.
Для исследования апоптоз индуцирующей активности рекомбинантных плазмидных ДНК использовали р53-дефектные опухолевые клеточные культуры человека А431 (эпидермальная карцинома) и С33А (карцинома шейки матки), в качестве нормальных клеток использовали культуру клеток MRC-5 (диплоидная культура легких эмбриона человека).
Для проведения трансфекции использовали реагент для трансфекции Lipofectamine™ LTX и реагент Plus (Invitrogen, USA). Процент апоптических клеток измеряли по методу, предложенному Афанасьевым [11] по наличию фрагментации ДНК через 48 ч после трансфекции. При этом фрагментированная в процессе апоптоза ДНК вымывается из клеток и при измерении сигнала флюоресценции PI на проточном цитометре апоптирующие клетки выпадают из области клеток с одинарным набором хромосом. Результаты исследования приведены в примере 1. Апоптоз индуцирующую активность сконструированных рекомбинатных плазмид также определяли по удельному содержанию жизнеспособных клеток в транфецированных опухолевых и нормальных культурах клеток с помощью счетчика клеток Scepter 2 (Milipore) (см. пример 2). Представленные результаты показывают, что сконструированные рекомбинантные плазмиды, экспрессирующие ген белка апоптина, индуцируют апоптоз использованных неопластических клеток, но не нормальных нетрансформированных клеток, что свидетельствует о селективности их воздействия на опухолевые клетки человека. Полученные результаты также демонстрируют, что рембинантная плазмидная ДНК pcDNA4-Apo-2NLS2, несущая ген апоптина с удвоенным сайтом NLS2, имеет апоптоз индуцирующую активность в отношении опухолевых клеток, достоверно превышающую таковую плазмиды pcDNA4-Apo экспрессирующей природный генетически немодифицированный ген белка апоптина.
Маркерные генетические признаки. Плазмидная ДНК pcDNA4-Apo-2NLS2 представляет собой эукариотическую плазмиду pcDNA4TOPO с инсерцией гена белка апоптина под цитомегаловирусный промотор, с удвоенным сайтом NLS2.
Репродуктивные свойства. Для наработки плазмидной ДНК pcDNA4-Apo-2NLS2 рекомендуется использование следующих штаммов E.Coli TOP10F', JM109 и INVaF' и протоколов производителя - Invitrogen.
Условия консервации и хранения. Хранить при -200С.
Патогенность для человека и лабораторных животных. Плазмидная ДНК pcDNA4-Apo-2NLS2 непатогенна.
Пример 1. Исследование апоптоз индуцирующей активности рекомбинантной плазмидной ДНК pcDNA4-Apo-2NLS2 при трансфекции опухолевых и нормальных клеток человека.
Проведено исследование апоптоз индуцирующей активности рекомбинантной плазмидной ДНК pcDNA4-Apo-2NLS2 в отношении опухолевых и нормальных клеток человека. С этой целью использовали р53-дефектные опухолевые клеточные культуры человека А431 (эпидермальная карцинома) и С33А (карцинома шейки матки), в качестве нормальных клеток использовали культуру клеток MRC-5 (диплоидная культура легких эмбриона человека).
Трансфекцию проводили с использованием реагента для трансфекции Lipofectamine™ LTX и реагента Plus (Invitrogen, USA). Трансфекцию проводили в шестилуночных культуральных планшетах с субконфлюентными монослоями опухолевых и нормальных клеток смесью соответствующих рекомбинантных плазмидных ДНК с реагентами для трансфекции в соответствии с рекомендацией производителя (2.5 мкг плазмидной ДНК+ липофектамин, 10 мкл + реагент Plus, 3.5 мкл) в 1 мл среды Opti-MEM (Invitrogen, USA). Процент апоптических клеток измеряли по методу, предложенному Афанасьевым [11] по наличию фрагментации ДНК через 48 ч после трансфекции. Клетки отбирали с культуральных планшет с помощью трипсина в 15 мл пробирки, центрифугировали при 1500 об/мин, отмывали в 1мл фосфатного буфера при 5000 об/мин 2-5мин. После чего клетки ресуспендировали в 300 мкл фосфатного буфера и добавляли 700 мкл 96% этилового спирта при -20°C. Фиксацию клеток проводили при -20°C в течение 30 мин. Перед измерением суспензии клеток отмывали 1 раз в фосфатном буфере и ресуспендировали в раствор для прокраски ДНК (2.5 мкг/мл пропидий йодида (PI), 0.5 мг/мл РНКазы, 1мл фосфатного буфера) на 30 мин при комнатной температуре. Меченые клетки измеряли при помощи проточного цитометра BioRad 200 по сигналу PI.
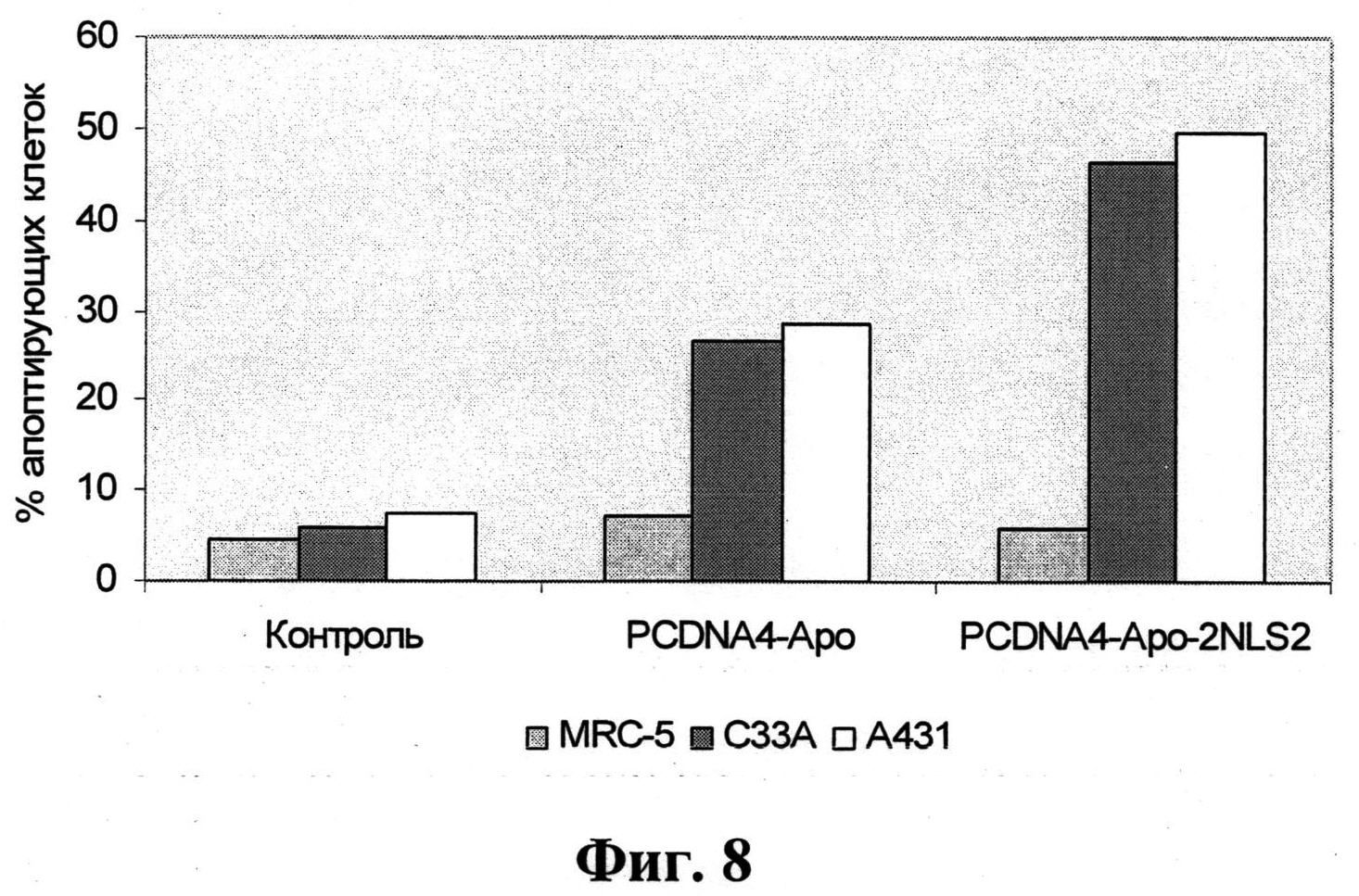
На фиг.8 приведена избирательная апоптоз индуцирующая активность рекомбинатных плазмид с инсерциями гена белка апоптина в отношении опухолевых клеток человека. Результаты, представленные на фиг.8, показывают, что рекомбинантная плазмидная ДНК pcDNA4-Apo-2NLS2 эффективно индуцирует апоптоз опухолевых клеток С33А и А431 (45-50% апоптирующих клеток) и не оказывает достоверного влияния на нормальные диплоидные клетки MRC-5. Плазмидная ДНК pcDNA4-Apo, несущая генетически не модифицированный ген белка апоптина, также достоверно индуцирует апоптоз опухолевых клеток, однако с существенно меньшей эффективностью (25-28% апоптирующих клеток) нежели pcDNA4-Apo-2NLS2. В качестве контроля использовали клетки трансфецированные плазмидой pcDNA4 без целевой инсерции.
Пример 2. Определение удельного содержания жизнеспособных клеток в трансфецированных рекомбинантными плазмидами опухолевых и нормальных культурах клеток с помощью счетчика клеток Scepter (Millipore).
Трансфекцию опухолевых и нормальных диплоидных клеток проводили, как указано в примере 1. Через 48 ч после трансфекции клетки отбирали с субстрата с помощью раствора Версена, отмывали центрифугированием при 1500 об/мин в 10 мл фосфатного буфера. После чего клетки ресуспендировали в 1 мл фосфатного буфера. Анализ проводили с использованием счетчика клеток Sceptertm 2 (Millipore) согласно инструкции производителя.
Результаты, приведенные в таблице 1, косвенно подтверждают высокую апоптоз индуцирующую активность плазмидной ДНК pcDNA4-Apo-2NLS2 в отношении используемых опухолевых клеток человека. Плазмидная ДНК pcDNA4-Apo, несущая последовательность природного гена белка апоптина, имеет существенно меньшую цитопатическую активность в отношении опухолевых клеток, чем pcDNA4-Apo-2NLS2, и также как и последняя не оказывает достоверного влияния на жизнеспособность диплоидных клеток MRC-5. Используемая в качестве контроля плазмидная ДНК pcDNA4 без целевой инсерции при проведении трансфекции не оказывает достоверного влияния на жизнеспособность ни опухолевых, ни диплоидных клеток.
Таблица 1
Процент жизнеспособных клеток в клеточных популяциях трансфецированных рекомбинантными плазмидами опухолевых и нормальных культур клеток.
|
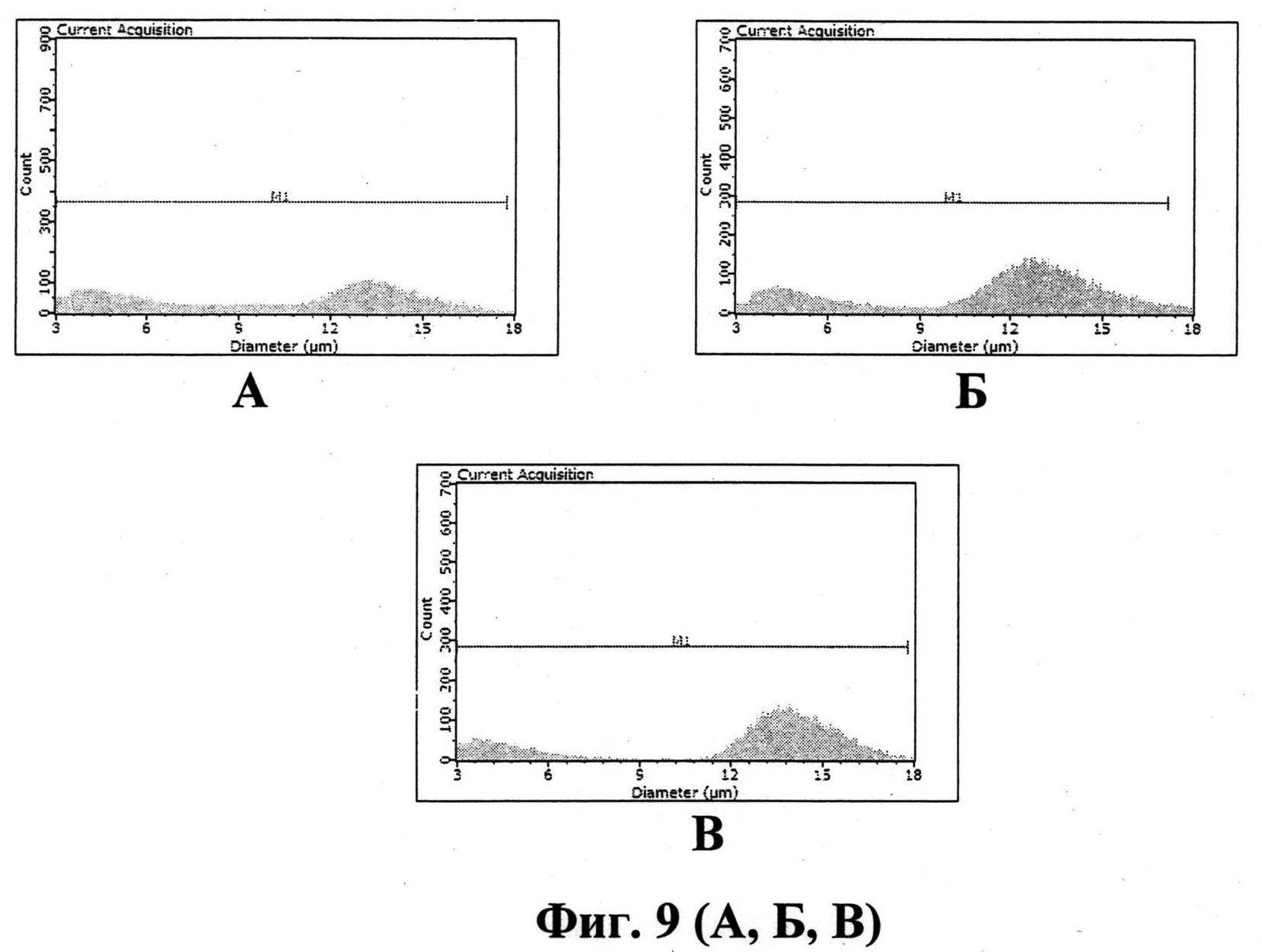
На фиг. 9 приведено распределение клеток С33А, определяемое цитометром Sceptertm 2, трансфецированных плазмидными ДНК pcDNA4-Apo-2NLS2 (А), pcDNA4-Apo (Б) и pcDNA4 (В).
Для иллюстрации (фиг. 9) воздействия рекомбинантных плазмид на жизнеспособность трансфецированных клеток представлены первичные данные подсчета клеток прибором Sceptertm 2 (опухолевые клетки С33А транфецированные плазмидами pcDNA4-Apo-2NLS2, pcDNA4-Apo и pcDNA4). Как видно, максимальное влияние оказывает плазмидная ДНК pcDNA4-Apo-2NLS2 (отмечается выраженное снижение доли жизнеспособных клеток диаметром 12-15 мкм и увеличение количества клеточного дебриса диаметром менее 11 мкм). Плазмида pcDNA4-Apo при трансфекции клеток С33А оказывает менее выраженное влияние на распределение клеток, определяемое цитометром Sceptertm 2, а при трансфекции клеток pcDNA4 достоверных отличий от контрольных клеток не выявляется.
Источники информации
1. Noteborn M.H., Todd D., Verschueren C.A., de Gauw H.W., Curran W.L., Veldkamp S., Douglas A.J., McNulty M.S., van der E.A., Koch G. A single chicken anemia virus protein induces apoptosis.// J Virol. - 1994. -V. 68. -P. 346-351.
2. Maddika S., Mendoza F.J., Hauff K., Zamzow C.R., Paranjothy T., Los M. Cancer-selective therapy of the future: apoptin and its mechanism of action.// Cancer Biol Ther. - 2006. -V. 5. -P. 10-19.
3. Noteborn M.H. Chicken anemia virus induced apoptosis: underlying molecular mechanisms.// Vet Microbiol. - 2004. -V. 98. -P. 89-94.
4. Heilman D.W., Teodoro J.G., Green M.R. Apoptin nucleocytoplasmic shuttling is required for cell type-specific localization, apoptosis, and recruitment of the anaphase-promoting complex/cyclosome to PML bodies.// J Virol. - 2006. -V. 80. -P. 7535-7545.
5. Poon I.K., Oro C., Dias M.M., Zhang J., Jans D.A. Apoptin nuclear accumulation is modulated by a CRM1-recognized nuclear export signal that is active in normal but not in tumor cells.// Cancer Res. - 2005. -V. 65. -P. 7059-7064.
6. Los M., Panigrahi S., Rashedi I., Mandal S., Stetefeld J., Essmann F., Schulze-Osthoff K. Apoptin, a tumor-selective killer.// Biochim Biophys Acta. - 2009. -V. 1793. -P. 1335-1342.
7. Toth K.,Wold W.S. Increasing the efficacy of oncolytic adenovirus vectors.// Viruses. - 2010. -V. 2. -P. 1844-1866.
8. Wu Y., Zhang X., Wang X., Wang L., Hu S., Liu X., Meng S. Apoptin enhances the oncolytic properties of Newcastle disease virus.// Intervirology. - 2012. -V. 55. -P. 276-286.
9. Hui Zhan, Jian-Song Wang, Hai-Feng Wang, Yi-Gang Zuo, Chun-Hui Wang, Ming-Xia Ding. Apoptin Induces Apoptosis in Human Bladder Cancer EJ and BIU-87 Cells.// Asian Pacific J Cancer Prev, - 2012. -V. 13. -P. 135-138 (прототип).
10. Li X., Liu Y., Wen Z., Li C., Lu H., Tian M., Jin K., Sun L., Gao P., Yang E., Xu X., Kan S., Wang Z., Wang Y., Jin N. Potent anti-tumor effects of a dual specific oncolytic adenovirus expressing apoptin in vitro and in vivo.// Molecular cancer. - 2010. -V. 9. -P. 10.
11. Afanas'ev V.N., Korol B.A., Mantsygin Yu.A. Flow cytometry and biochemical analysis of DNA degradation characteristic of two types of cell death. // FEBS Lett. - 1986. № 194. - P. 347-350.
Приложение
<110> Федеральное бюджетное учреждение науки «Государственный научный центр вирусологии и биотехнологии ′′Вектор′′» (ФБУН ГНЦ ВБ ′′Вектор′′)
<120> Рекомбинантная плазмидная ДНК pcDNA4-Apo-2NLS2, несущая синтетический ген белка апоптина, индуцирующего р53 независимый апоптоз опухолевых клеток
<160> 1
<210> SEQ ID NO1
<211> 393
<212> DNA
<213> Chicken anemia virus (вирус анемии цыплят)
<400> 1 Кодирующая область синтетического гена белка апоптина
ATGA ACGCTCTCCA AGAAGATACT CCACCCGGAC CATCAACGGT GTTCAGGCCA CCAACAAGTT CACGGCCGTT GGAAACCCCT CACTGCAGAG AGATCCGGAT TGGTATCGCT GGAATTACAA ТСАСТСТАТС GCTGTGTGGC TGCGCGAATG CTCGCGCTCC CACGCTAAGA TCTGCAACTG CGGACAATTC AGAAAGCACT GGTTTCAAGA ATGTGCCGGA CTTGAGGACC GATCAACCCA AGCCTCCCTC GAAGAAGCGA TCCTGCGACC CCTCCGAGTA CAGGGTAAGC GAGCTAAAAG AAAGCTTGAT ТАССАСТАСТ CCCAGCCGAC CCCGAACCGC AAGAAGGTGT ATAAGACTGC GAACCGCAAG AAGGTGTATA AGACTGTAA
Рекомбинантная плазмидная ДНК pcDNA4-Apo-2NLS2, несущая синтетический ген белка апоптина с удвоенным сайтом NLS2 и предназначенная для экспрессии указанного белка апоптина, избирательно индуцирующего р53 независимый апоптоз неопластических опухолевых клеток человека, с нуклеотидной последовательностью SEQ ID NO1 размером 393 п.н., представленной на фиг. 4, имеющая молекулярную массу 5,13 мДа, размер 7896 п.н. и содержащая в соответствии с физической и генетической картой, приведенной на фиг. 5:- оперон, размером 1071 п.н., состоящий из ранне-позднего промотора CMV, лидерного фрагмента, последовательности пенетрирующего пептида белка ТАТ ВИЧ, гена апоптина вируса анемии цыплят и последовательности эпитопа Flag;- промотор CMV 232-819 п.н.;- Т7 промотор 863-882 п.н.;- Xpres праймер 1112-1130 п.н.;- ТОРО сайт клонирования 1184 п.н.;- генетические маркеры: ген устойчивости Zeocin; ген b-лактамазы, определяющий устойчивость к ампициллину;- уникальные сайты рестрикции: ClaI (1918); XbaI (2619); BamHI (2993); SalI (3001); BstWI (3425).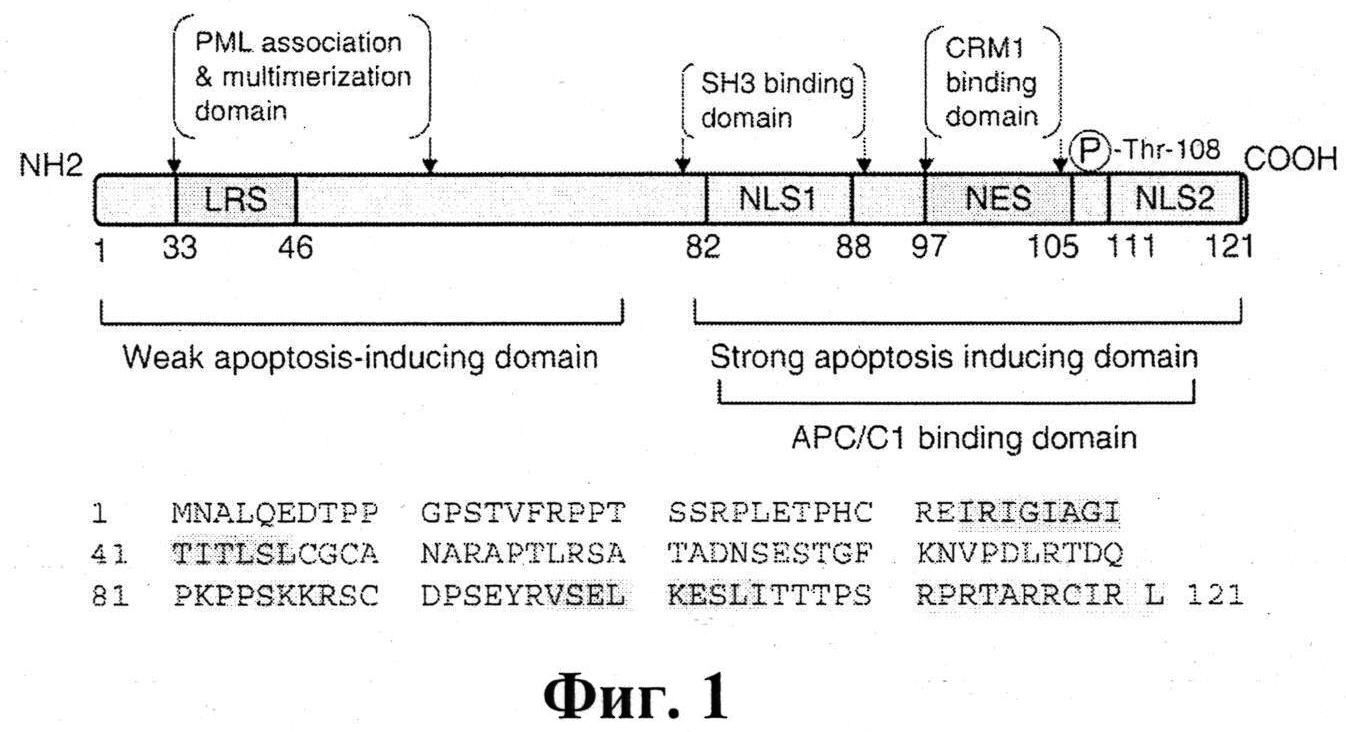



Рекомбинантные плазмидные днк pqe-60-tnfr-crmb-ind-67 и pfastbac1-g2r-dsecret, содержащие фрагмент генома вируса натуральной оспы, кодирующий фактор некроза опухолей связывающий домен белка crmb и штамм бакуловируса bv/g2r-dsecret, продуцирующий секретируемый фно-связывающий белок crmb вируса натуральной оспы с делетированным secret-доменом
Набор олигодезоксирибонуклеотидных праймеров и флуоресцентно меченых зондов для идентификации рнк коронавирусов видов 229е, nl63, ос43, hku1 методом гибридизационно-флуоресцентной обратно-транскриптазной полимеразной цепной реакции
Способ получения слабоположительной контрольной сыворотки, содержащей ad и ay субтипы hbsag
Ингибитор репродукции вируса гриппа а на основе экстракта базидиального гриба phallus impudicus
Ингибитор репродукции вируса гриппа а на основе экстракта базидиального гриба laetiporus sulphureus
Противовирусное средство на основе штамма нематофагового гриба duddingtonia flagrans f-882
Биологический микрочип на основе диэлектрофореза, система для экспресс-идентификации вирусов и способ экспресс-идентификации вирусов с их использованием
Штамм вируса гриппа a/teal/chany/7119/08/ h15n4-субтипа для получения антигенсодержащего препарата и поликлональной сыворотки для диагностических целей
Набор олигонуклеотидных праймеров и флуоресцентно-меченых зондов для субтипоспецифичной идентификации рнк вируса денге на основе мультиплексной пцр в реальном времени
Набор олигонуклеотидных праймеров и флуоресцентно-меченых зондов для видоспецифичной экспресс-идентификации вируса эбола-судан методом полимеразной цепной реакции
Рекомбинантные плазмидные днк pqe-60-tnfr-crmb-ind-67 и pfastbac1-g2r-dsecret, содержащие фрагмент генома вируса натуральной оспы, кодирующий фактор некроза опухолей связывающий домен белка crmb и штамм бакуловируса bv/g2r-dsecret, продуцирующий секретируемый фно-связывающий белок crmb вируса натуральной оспы с делетированным secret-доменом
Набор олигодезоксирибонуклеотидных праймеров и флуоресцентно меченых зондов для идентификации рнк коронавирусов видов 229е, nl63, ос43, hku1 методом гибридизационно-флуоресцентной обратно-транскриптазной полимеразной цепной реакции
Способ получения слабоположительной контрольной сыворотки, содержащей ad и ay субтипы hbsag
Ингибитор репродукции вируса гриппа а на основе экстракта базидиального гриба phallus impudicus
Ингибитор репродукции вируса гриппа а на основе экстракта базидиального гриба laetiporus sulphureus
Противовирусное средство на основе штамма нематофагового гриба duddingtonia flagrans f-882
Биологический микрочип на основе диэлектрофореза, система для экспресс-идентификации вирусов и способ экспресс-идентификации вирусов с их использованием
Штамм вируса гриппа a/teal/chany/7119/08/ h15n4-субтипа для получения антигенсодержащего препарата и поликлональной сыворотки для диагностических целей
Набор олигонуклеотидных праймеров и флуоресцентно-меченых зондов для субтипоспецифичной идентификации рнк вируса денге на основе мультиплексной пцр в реальном времени
Набор олигонуклеотидных праймеров и флуоресцентно-меченых зондов для видоспецифичной экспресс-идентификации вируса эбола-судан методом полимеразной цепной реакции

